

If you haven’t been following along with Julie Reynolds‘ delightful series on phenotypic plasticity, I recommend catching up on that first. Julie shared some great real world examples, but not everyone has the opportunity to study overwintering insects like she does. So I thought I’d give you a hands-on example, albeit a simulated one. I’ve introduced my Quandary Den before. Briefly, players have to ‘zap’ or ‘tag’ robots for points, but the players have to evolve their gameplay approach. The versions I’ve shared before do not have any capacity for phenotypic plasticity, but we can change that.
We can actually keep the genotype (a series of letters) and the phenotype (a series of moves in the game) the same and just change how we map the one to the other. Previously, we read the genotype sequence as a sequential series of moves – go left, go down, zap right, and so on. As a result, the phenotype will be the same regardless of where the robots are; if we put the robots in new spots but keep the players’ genotype the same, the players will still make the same moves and possibly not do very well. Some examples are shown below; even just small shifts in the robot positions renders a previously perfect player genotype woefully ineffective.
To introduce more flexibility, we can instead interpret the genotype as a strategy for various game scenarios. We’ll define the scenarios as what the player can ‘see’ in each direction: up, down, left and right. So if the player is in the lower left corner with a robot above them, the scenario is robot, wall, wall, nothing. We can list every possible scenario and put them in some order of our choosing. Then we can read the genome as indicating in order which action to take in each of those scenarios. Once the game starts, we figure out which scenario the player is starting in, look up the action for that scenario and perform it, then recalculate the scenario and repeat. This way, the same genotype can result in a variety of phenotypes depending on where the robots are. Below are examples; the genotype is the same and so the strategy is the same, but the specific actions at each moment depend on the scenario or context, allowing the player to be successful even when the robots are moved.
I’ve implemented this version for you to experiment with. You can choose between the original deterministic genotype/phenotype match and the new contextual one. You can also change the other variables we’ve looked at before, like whether neutral mutations can accumulate or whether there can be multiple players. You can also decide whether the robots start in the usual place or a random one, and whether their position changes after each generation or not.
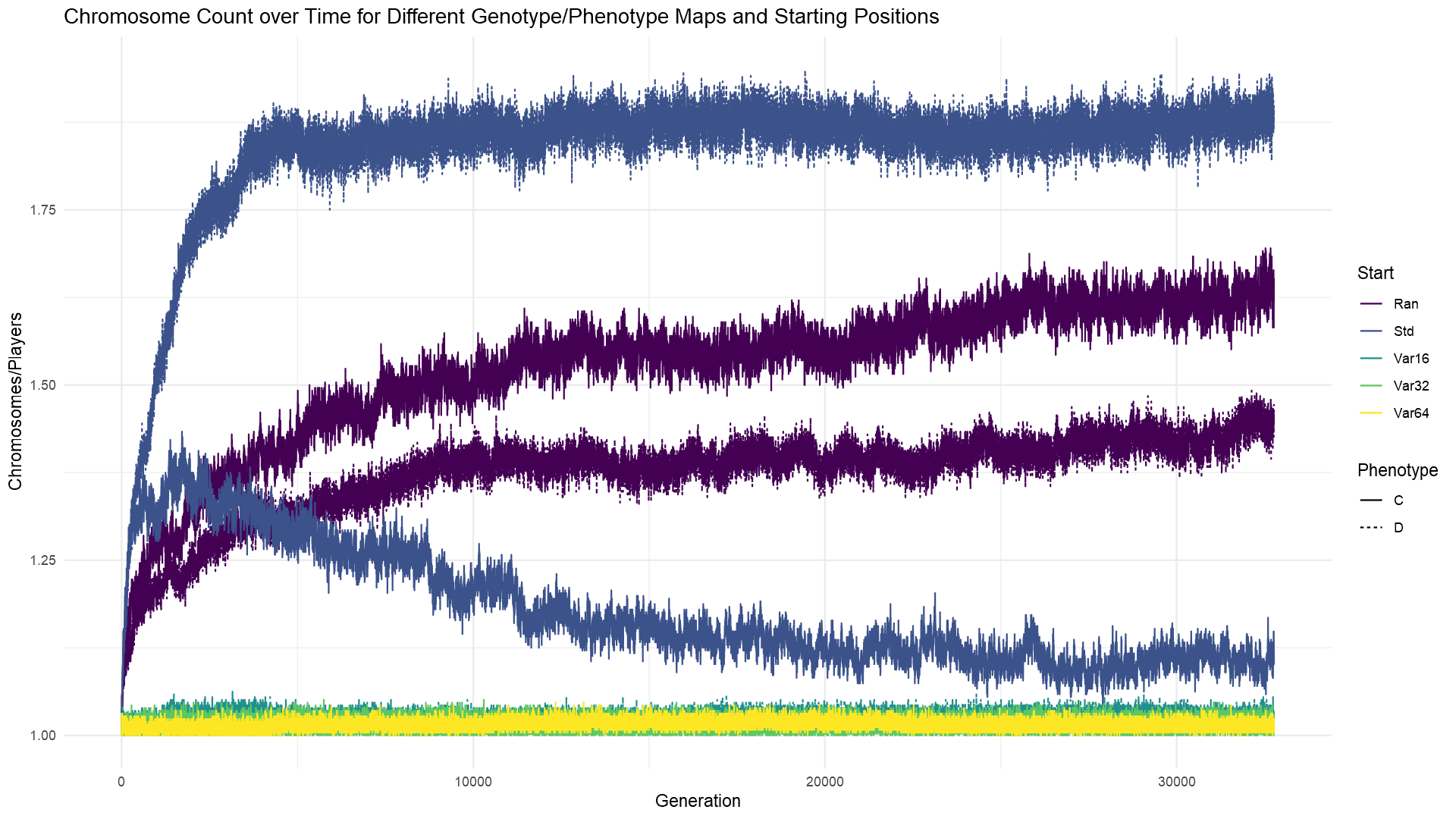
For some initial exploration, I’ve run a few experiments and plotted the fitness results in the chart at the top of the post. Dashed lines are for the deterministic genotype/phenotype map (‘D’) and solid lines are for the contextual one (‘C’). Standard starting positions are dark blue (‘Std’); random but fixed between generation starting positions are purple (‘Ran’); random and varying between generation starting positions are in light blue, green and yellow lines, averaging over 16, 32, or 64 different positions each generation (‘Var16’, ‘Var32’, ‘Var64’). I averaged over multiple starting positions each generation when the positions were random and varying in order to smooth out any impact of a particularly good or bad random arrangement. And I was curious if the number of different positions would matter; the effect seems small, but having to solve more positions seems to be harder.
There are a few other things we can note from these results. First, it seems that my standard starting positions (‘Std’) present a particularly easy challenge compared to random fixed positions (‘Ran’). It also looks like the deterministic approach has the advantage when the starting positions don’t change, getting to higher fitness faster; I’ll elaborate on why I think that is in a bit. However, the situation flips when the positions change each generation; in that case, a contextual strategy can get better results more quickly. And therein lies at least part of an answer for why it is worthwhile for organisms to have some phenotypic flexibility: dealing with changing, unpredictable conditions. For the curious, you can also check out the average number of chromosomes that evolved in each experiment; feel free to share your observations in the comments.
In addition to illustrating phenotype plasticity, this change brings up some other aspects of evolutionary biology. First is differences in fitness landscapes. The evolutionary process tends to select for populations with higher average fitness, which can be visualized metaphorically like climbing a hill with the top corresponding to the highest possible fitness. That hill image prompts the further metaphor of a fitness landscape. The simplest fitness landscape has a single, tall hill surrounding be plains that slope away from the hill in all directions. But just as with actual terrain, other fitness landscapes are possible, ones with multiple hills of varying heights and steepness. Changing to the contextual genotype/phenotype map results in a more varied fitness landscape, with hills that have peaks that are not as high as other hills’ but that have valleys in between. Consequently, if the player evolved a genotype corresponding to the peak of one of those low hills, mutations with lower fitness would have to accumulate before a higher hill could be climbed. By contrast, with the deterministic genotype/phenotype map the evolutionary process can keep taking steps upward (or sidewise, for neutral mutations) and ultimately get to the highest possible peak. This may explain the advantage the deterministic version has in the fixed starting position experiments.
The other evolutionary topic raised by changing genotype/phenotype map is the consistency of selection. When we keep the robots in the same place throughout the evolutionary process, we give the players an opportunity to learn those specific locations. With the contextual genotype/phenotype map, it makes more sense to move the robots around to see if we can evolve more general strategies that apply to a wide range of positions. (Although in hindsight, there are solutions in the deterministic version that would apply to multiple robot positions as well.) In a biological context, this would correspond to changing environments and changing selection pressures. Take for example the celebrated peppered moths; after increases in air pollution from the Industrial Revolution, darker moths survived better because they blended in with now-darker tree trunks, but subsequent clean air initiatives reduced pollution and the lighter moths became the most common again. Or consider the recent story about elephant populations without tusks, having evolved under selection pressure from poaching; it is expected that if poaching is brought under control, tusked elephants will rebound.
Fluctuating environmental conditions can lead to long-term consistency in populations even if there are short-term changes. Thus lasting population change is more likely if there is consistent selection in a particular ‘direction.’ This can happen when new niches are created or open up, such as when the atmosphere became oxygen-rich or when dinosaurs went extinct. This may partly explain why the fossil record shows periods of little or no change and periods of rapid change. When the environment is on average the same over time, the organisms living in it will tend to be the same as well. But when there is a persistent change in the environment, there are opportunities for new organisms.
Andy has worn many hats in his life. He knows this is a dreadfully clichéd notion, but since it is also literally true he uses it anyway. Among his current metaphorical hats: husband of one wife, father of two teenagers, reader of science fiction and science fact, enthusiast of contemporary symphonic music, and chief science officer. Previous metaphorical hats include: comp bio postdoc, molecular biology grad student, InterVarsity chapter president (that one came with a literal hat), music store clerk, house painter, and mosquito trapper. Among his more unique literal hats: British bobby, captain’s hats (of varying levels of authenticity) of several specific vessels, a deerstalker from 221B Baker St, and a railroad engineer’s cap. His monthly Science in Review is drawn from his weekly Science Corner posts — Wednesdays, 8am (Eastern) on the Emerging Scholars Network Blog. His book Faith across the Multiverse is available from Hendrickson.
Sir, macro-evolutions have already been proved to be absolutely impossible for many many reasons such as these:
1. The notion of species are not strictly defined.
2. The two kinds of “Darwin’s dilemmas/contradictions”:
A. No sudden Cambrian explosion by random mutations can happen, probability-wise.
B. The most numerous species are the least-evolved, least-fittest-to-survive, lowliest creatures.
3. Popper’s dilemma/contradictions:
It is impossible probability-wise that both controlling central nervous systems and peripheral motor/sensory organs to be controlled by them, inside the bodies of individuals of an old species, macro-evolve at the same time, which, however, must have happened according to macro-evolutionism:
For example, the wings-controlling central nervous systems of reptiles turning into birds can never logically evolve before the wings. But the opposite is also impossible; birds can not fly in controlled ways without such nervous systems to control their wings, etc..
4. See my criticisms against the notions of chaos and complex systems and emergences incl. macro-evolutions at @koitiluv1842 on Twitter.com.
5. Macro-evolutionism even lacks any theoretical working hypotheses to explain the macro-evolution processes with, let alone any empirical methods or data to verify them with.
6. Self-organizations produce only patterns like AAAA BB CCCC DDD, but no meaningful info like in “The cat caught the mouse.”
7. See Werner Gitt: “Without Excuse” or “In The Beginning Was Information” for other info-theoretical macro-evolutionism refutation.
8. See also Jonathan Wells: “Icons of Evolution” for the exposures of blatant lies in the drawings that have been used to support macro-evolutionism.
9. See also: https://www.icr.org/article/mathematical-impossibility-evolution/
I admit that Wisdom or Sirach has descriptions of God’s artificial breeds-improvements of land animals/birds into water animals/birds and vice versa, but they were no macro-evolutions.
Let’s take a look at this points, shall we?
1. True, there is not a single, unambiguous definition of species. There is the biological concept of a species, but that can only be applied to living organisms and still results in some ambiguities (e.g. ring species). Criteria applied to fossils are not guaranteed to align with the biological concept in all cases. But why is that a problem for macroevolution? Isn’t that to be expect from a model of common descent in which populations are constantly changing and divergence has no particular endpoint? What alternative model is more consistent with the observed complications of defining species?
2. a. Analysis of the phylogenetic relationships of arthropods, the most diverse group of animals, shows that diversification in the Cambrian required a mutation rate consistent with observed values in the present. https://www.sciencedirect.com/science/article/pii/S0960982213009160
2. b. I assume by “most numerous species” you mean “species with the largest population sizes” in which case we are talking about bacteria. Arguably these are the most evolved organisms, in the sense that they have gone through the largest number of generations since their divergence from all their other cousins. And the very fact that they are so numerous speaks to a high fitness rather than being the least fit to survive, not to mention that they can live longer and in more extreme conditions than most other organisms.
3. To the specific example, bird wings did not emerge all at once but evolved from theropod limbs which already had a central nervous system which could control those limbs. The plasticity of nervous systems suggests that the ability to learn specific movements, such as those to achieve flight, are more dependent on the appropriate skeleton and musculature than the central nervous system. And fossil evidence suggests those limbs had a wing-like morphology prior to flight. So I don’t see what the challenge is here to macroevolution.
4. Do you have a more direct link? I scrolled through your Twitter feed and did not see a relevant article or discussion in the recent past.
5. One doesn’t need to search hard to find numerous examples of mathematical models for macroevolution. So I don’t really understand this claim. Are there specific macroevolutionary processes you see as under-developed?
6. & 7. Earlier in this series I showed an experiment which demonstrates how meaningful information can arise from an evolutionary process. The chart at the bottom of this post shows how information about the location of the robot has accumulated in the players’ “genome” via an evolutionary process.
8. Yes, Ernst Haeckel’s drawings are still shown in many textbooks, and yes, they are inaccurate in significant ways. At the same time, current understanding of the genetics and processes of development still points to common ancestry, even if ontogeny does not recapitulate phylogeny as strongly as Haeckel asserted. We could get rid of those drawings from all educational materials, and I have no objections to such a change, and it would not change the strength of the evidence for common descent and macroevolution one bit.
9. That article asserts that systems with more components are less likely, but again I’ve shown in this blog series that the opposite is true. Or just consider the final chart in the post above. Further, the probability calculations in that article only works out the probability of one specific combination of 200 mutations. But we really need to know is the probability of the entire class of 200-part systems which perform some advantageous function.
Thank you for your reply, but I disagree with your points. In a nutshell, you resorted to kinds of illogic.
For example:
1. You say that bacteria, not us humans, are “THE most evolved” species.
2. By “wings-controlling central nervous systems,” I of course meant such brain area(s). Do you have any fossils-based evidence(s) that what you meant by “wings-controlling central nervous systems” were as large and functionally sophisticated as such brain area(s)?
3. Here’s one of my many chaos and complex-systems criticisms’ outlines:
https://twitter.com/koitiluv1842/status/1457320230758195200
Do you agree to chaos/chaotic “theory” that most part of this world is a chaos or “chaosmos” = crossbreed between chaos and cosmos/order = complex system, which allows the un-defined/contents-less “emergences” including macro-evolutions? Namely, do you assume that the orderly God Almighty is NOT ruling/controlling all events orderlily, not chaotically, and therefore, also that there will NOT be any orderly/righteous Judgement against the evil ones’ atrocities based on their free-will granted to them by Loving God? Do you advocate that, after all, the world is chaotic, so, let’s just lead chaotic/wild/God-unfearing, hedonic lives?
In the first place, you started out by denying the Biblical truth below:
“Life Application Study Bible: Personal Edition with New Living Translation, Second Edition” (Tyndale House Publishing, 2007),
Genesis 1:25 — “God made all sorts of wild animals, livestock, and small animals, each able to produce offspring of the same kind.” Similar verses in Genesis 1:12 and 1:21 states similar things about plants and water-animals and birds.
(Pls see also the below website of Herr Hutschi = Hutschenreuther, where my math theology is posted, if possible.
Note: Herr Hutschi is an anti-Christ. He is only interested in the relation between science and religiion.)
God is in control/charge of everything including all the stochastic = only-seemingly-chaotic = intrinsically-pseudorandom processes. Thus, there are no “chaos,” no “complex systems” on “the edge of chaos” or as crossbreeds between “chaos” and order.
Here are two of the reasons why:
1. The universality of LLNs (= the strong, weak, and complete laws of large numbers) = the criticisms against the “exceptions to LLNs”:
https://twitter.com/koitiluv1842/status/1472172686638206976
2. The non-existence of any un-computable or truly-/NON-PSEUDO-“random/chaotic-numbers” (Excuse me the Japanese posts, and that they are only outlines of my proofs):
https://twitter.com/koitiluv1842/status/1472510570104778755
https://twitter.com/koitiluv1842/status/1472526559278157830
(The above are my posts concerning the links of Prof. Junpei Tsuji’s blogs.)
On 5.:
Listen, Dr./Prof. Walsh: I was not born yesterday; I majored in the German language to minor in the Marxist philosophy including the Marxist politico-economics in order that I could pursue/explore thoroughly “the wonderful Marxist atheist world view,” searching and searching for decades for the perfect concrete theory of the concrete mechanism of macro-evolutions through natural selections by random mutations that are based on “dialectical” or some other “physical” principles of the atheist, God-less self-motions/self-developments/self-evolutions of matters in general and of those of living things in particular; but in vain at all !
1. As I repeated in my previous replies, undefined emergences or self-organizations fail to work for macro-evolutionism.
2. Emergences “defined” as some “chaotic” motions of complex systems also fail to work as shown in one or two of my previous replies.
3. The below book, too, confess that the authors fail to know or understand the crucial part of the concrete method to resolve Darwin’s dilemmas:
https://www.amazon.co.jp/-/en/Marc-W-Kirschner/dp/0300108656/ref=sr_1_1?crid=X55084FOTZV6&keywords=Plausibility+of+Life+Resolving+Darwin%E2%80%99s+Dilemma&qid=1640393812&s=english-books&sprefix=plausibility+of+life+resolving+darwin+s+dilemma%2Cenglish-books%2C156&sr=1-1
4. The following book by Gregory J. Chaitin does not present any successful concrete macro-evolution mechanism theory, either, especially because I have refuted his DM (= diagonal method), which pls see in one or a few of my previous Tweets posts or other Tweets of mine:
https://www.amazon.co.jp/Proving-Darwin-Making-Biology-Mathematical/dp/0375423141
When you admit the macro-evolutionism for matters in general (= macro-evolutions of coacervates or some RNA-slimes into protozoa) and for living things in general, you are denying the existence and intervening works of The All-Knowing, All-Powerful Creator = Controller = Vindicator = Judge = Redeemer/Savior who knew and know all the initial and interim conditions for all the states of all the elementary particles in the whole universe that He Created about 6,000 years ago (according to The Book of Jubilees = Leptogenesis = Lesser Genesis).
In other words:
Do you intend to assert that God leaves us to random/chaotic/wild/stochastic fates by throwing dice?
To be (a) true Christian(s) or not to be one(s), that is the question for you and followers of “theories” like yours!
Notes on the minor errors in Lee Strobel’s and Werner Gitt’s books:
1. Both of them approves Cantor’s and Goedel’s and others’ DM (= diagonal methods) ⇔ Einstein’s TR (= theories of relativity), for whose equivalency and for whose disproof, pls see my one or a few of my previous Tweet links.
2. Strobel goes further to say, following Karl Popper’s falsifiability “theory” based on Geodel’s-DM (= Incompleteness Anti-Theorem based on his Diagonal Lemma) and TR, that Christian theology were a truth because it were “falsifiable,” and that macro-evolutionism were a falsehood because it were “un-falsifiable”.
3. Strobel praises his interviewee Meyer’s groundless attack-attempt on Cartesian theology (although Descartes calls himself a Christian “philosopher”.)
4. Strobel lauds one of his interviewee’s “theology” that God were not The Infinite/Unlimited/Absolute because infinity could not be a mathematical concept because infinity minus infinity is indefinite; but zero is a mathematical concept although zero divided by zero is indefinite.
5. Strobel believes in one of his interviewee’s mis-interpretations of Michel Nostradamus’s “Proph’eties/Prophecies” (my criticisms against which I omit here.)
TR is false. ⇔ No “multiverse” can exist.
Also, pls see:
https://twitter.com/koitiluv1842/status/1474579184890806273
You must admit that the Darwinist-Marxist-Einsteinian-Goedelian-atheism or -deism/pantheism has been annihilated now. (Did you know Darwin and Marx was good friends? Darwin Museum has been exhibiting the copy of “The Capital” that Marx sent to Darwin.)
You seem to have ignored and dismissed my previous posts on my disproofs of all the tries at theories of chaos, of complex systems on the edge of chaos or between chaos and order, of emergences inside complex systems, of macro-evolutions as emergences (namely, of the tries such as the ones in the below books): disproofs by my Chaitin’s/etc.-DMd (= diagonal-methods disproofs) Tweets and by my exposure of the authors’ confessions that they also fail to know or understand any concrete macro-evolution mechanisms or any concrete method of resolutions of Darwin’s dilemmas.
https://www.amazon.co.jp/-/en/Gregory-Chaitin/dp/0375423141/ref=pd_ybh_a_1/357-0422962-2037833?pd_rd_w=eylBw&pf_rd_p=c4d2dacb-1bb4-426c-bb91-4cdb0fc95702&pf_rd_r=G09FFP5RR7CZ2CAFWCTE&pd_rd_r=71905f83-40fa-43ac-9bfa-ef194e2b248e&pd_rd_wg=KeMQ2&pd_rd_i=0375423141&psc=1
https://www.amazon.co.jp/-/en/Marc-W-Kirschner/dp/0300108656/ref=pd_ybh_a_2/357-0422962-2037833?pd_rd_w=eylBw&pf_rd_p=c4d2dacb-1bb4-426c-bb91-4cdb0fc95702&pf_rd_r=G09FFP5RR7CZ2CAFWCTE&pd_rd_r=71905f83-40fa-43ac-9bfa-ef194e2b248e&pd_rd_wg=KeMQ2&pd_rd_i=0300108656&psc=1
You have been denying the existence of The Unique, All-Knowing, All-Powerful/Infinite/Unlimited/Absolute Creator = Controller = Vindicator = Judge = Redeemer/Savior who knew and knows all the initial and interim conditions for all the states of all the elementary particles in the whole finite (= limited but open/not-closed) universe that He Created six-thousand years ago (according to The Book of Jubilee = Leptogenesis = Lesser Genesis) but who never leaves us to any random/chaotic/wild/stochastic fates by “throwing dice”: a fact that even Einstein wanted to prove but had got temporarily “counter-criticized” by the use of his own TR. (Now I have counter-counter-criticized that folly.)
Pls upload/show my previous posts to you on this website.
The other tweets of mine proved the nonexistence of all other kinds of chaos attractors: the sine qua non for the chaos “theory”.
My other tweets also disproved DM (= diagonal methods) like the DM of Gregory J. Chaitin in this book = “Proving Darwin: Making Biology Mathematical”:
https://www.amazon.co.jp/-/en/Gregory-Chaitin/dp/0375423141/ref=pd_ybh_a_2/357-0422962-2037833?pd_rd_w=RLHjA&pf_rd_p=c4d2dacb-1bb4-426c-bb91-4cdb0fc95702&pf_rd_r=A3J29KA7PP2YEBKZVGB8&pd_rd_r=06cd8d0d-6200-43e7-b83b-b47bffff7510&pd_rd_wg=0AeGo&pd_rd_i=0375423141&psc=1
The authors of the below book “Plausibility of Life: Resolving Darwin’s Dilemma” confess that even they actually fail to know any concrete mech of macro-evolutions, namely, any concrete way to resolve Darwin’s dilemmas:
https://www.amazon.co.jp/-/en/Marc-W-Kirschner/dp/0300108656/ref=pd_ybh_a_1/357-0422962-2037833?pd_rd_w=RLHjA&pf_rd_p=c4d2dacb-1bb4-426c-bb91-4cdb0fc95702&pf_rd_r=A3J29KA7PP2YEBKZVGB8&pd_rd_r=06cd8d0d-6200-43e7-b83b-b47bffff7510&pd_rd_wg=0AeGo&pd_rd_i=0300108656&psc=1
No results/fruits were gained by my decades-long hard searches for any perfect mathematico-physical theory on the concrete mech of macro-evolutions through natural selections by random mutations on the basis of “the dialectics” or some other “physical” principles.
There exists the unique, All-Knowing, All-Powerful/Infinite/Unlimited/Absolute Creator = Controller = Vindicator = Judge = Redeemer/Savior who has known all the initial and interim conditions for all the states of all the elementary particles in this finite (= limited but open/not-closed) universe that He Created and who never leaves us to stochastic/random/chaotic/wild fates because He “does not throwing dice,” as even Einstein wanted prove but failed to do so.
A couple of my other criticisms against chaos:
1. https://twitter.com/koitiluv1842/status/1471690213777088513
2. https://twitter.com/koitiluv1842/status/1464215651954819077
3. https://twitter.com/koitiluv1842/status/1469507084467191809
Thank you for providing additional links on the topic of chaos and emergence.
You posted a lot of material in the replies above. They over a holiday which is why I did not get to them right away, and it has taken some time to be able to come back to it all. The single largest theme seems to this mathematical one, so perhaps it will be most fruitful to start there.
I can understand why one would object on theological grounds to chaos as a substantial feature of creation. After all, the opening verses of the Bible are about God bringing order to the world in contrast to chaos. And while I’m reasonably certain I did not follow all of the technical details of your mathematical points (I am a biologist by training, not a mathematician), I did get a sense that you object to the characterization of the relevant math as random and disordered and chaotic when in fact it exhibits structure and order. If I have understood you correctly, then perhaps we agree that chaos theory can be an unhelpful name and that too much is made of the sensitivity to initial conditions while the stability implied by attractors is underappreciated.
As it happens, I have written about chaos theory and theology in my book Faith across the Multiverse. (https://www.amazon.co.jp/-/en/Andy-Walsh/dp/1683070763/) Perhaps that can provide a basis for further discussion.
On your 8.:
Not only Haeckel’s, but all the evolutionist drawings are only imaginations, as J. Wells pointed out.
Thank you very much for your reply! I am relieved to know that you had not ignore all those above replies of mine.
Anyway, no chaos means: no “edge of chaos,” no “complex system on the edge of chaos,” no “emergence as a behaviour of a complex system,” and no “macro-evolution regarded as a kind of emergence”. Which further means that the notion of macro-evolution has now lost all mathematical bases/grounds.
In fact, the “emergence” notion has only a subjectivistic definition of “an un-expected behavior of a complex system” = no objective, mathematical definition.
Last but not the least: I have ordered your book today. Thank you for letting me know of it. I hope I will survive this severe winter until the day(s) when we can discuss it.
Thank you for giving my book a try. In the interim, if I may ask a question, how do you describe the output of the logistic map as r approaches and then exceeds the value of ~ 3.56995? Do you agree that there is something quantitatively different about the output from the logistic map for r > 3.56995 than when r < 3, or do you reject the need to make such a distinction? Or is your point that mathematical models which exhibit such behavior have no correspondence to physical systems?
Excuse me my ignorance, but as you can see either in the link(s) in my previous reply/replies to you or in my other recent Twitter tweet(s) @koitiluv1842, I have only principle-wise disproved the exist-ability of the chaos attractors (as VECTOR fields) defined by logistic mappings etc. on no-front-and-back-sided/no-direction-ed/”un-orientable”/NO-VECTORS-allowing Moebius bands, because such “attractors” are self-contradictory notions; I have never bothered to study any detailed actual values-computations about them.
I am sure that you accept Cartesian/Newtonian/Riemannian dualist-physicalist/materialist philosophy=theology assuming both natural/”physical” “matters” and supernatural/spiritual “matters” to be handled by the Riemannian neo-psychophysics. That is because I am also certain that you acknowledge the justifiability of Lenin’s definition of objective/physical “matters/objects” as “objective entities which exist relatively independently from your mind but whose objective existence and natures you can cognize/presume by objective/empirical sense-data and consistent (= non-paraconsistent/inconsistent/dialectical/Marxist-Leninist) contents-logic/Inhaltslogik.
Then, you surely admit also that
1. The future philosophy=theology must rely on the dynamical-systems-theory, specially, elementary-catastrophy theory = ECT with combinations of EC’s like the double-cusps. (As Ren’e Thom, a modern Cartesian/Newtonian, correctly pointed out, all the concepts must be geometrical-ized.)
and
2. Math is the abstracted physics.
Thus, you must deny all the unreal/non-mathematico-physical/non-dynamical-theoretical notions like the twisted products with the (n+1/2)-times-twists = with no front/outer and/or back/inner side(s).
A bi-product of the above:
Heawood Conjecture has no exceptions for such twisted products like the non-existent Klein-bottle.
(Also, other presumed “exceptions” were negated later.)
Heawood Conjecture is, therefore, a universal theorem.
Thus, it makes the shortest supportive proof of the four-color theorem.
https://mathworld.wolfram.com/HeawoodConjecture.html
Thom meant, the meanings/semantics/images of all the concepts must get represented as catastrophe-theoretic n-D hyper-figures in motion.
For the equivalencies between the existences of logistic mappings, of period-doubling cascades, of Feigenbaum tree, and of Moebius band, please Google.
For my criticisms against them, please see my Twitter tweets.
I’m sorry, but I don’t see how any of these replies address my question. So let me try again. I can see dynamical systems exhibit period doubling with my eyes and verify it analytically. I’ve seen dynamical systems exhibit sensitivity to initial conditions. So are you saying that you’ve proved something counter to what I have observed, and that dynamical systems don’t have these properties at all? Or are you saying that they only have these properties in the realm of abstract mathematics, but no actual physical system can have these properties?
I regret to inform you that, in your previous reply, you missed my very most weighty point: NO chaos in any forms can and could exist mathematico-physically, from the beginning of the world: before and during and after the Creation of this “material” and spiritual universe. Which means, The Creation was the creatio ex deo = a transformation of an order/orderliness of the perfect God as a Spirit (= the then-unique/only, spiritual/supernatural “matter”) into another order/orderliness of the perfect, “material” and spiritual/supernatural universe before the falls of some angels-turned-evil-spirits (including Satan, the prince of evil spirits) and of the first-generation snake and of the first-generation humans.
When I wrote above about Genesis 1 discussing God bringing order in contrast to chaos, I was referring to the term tohu wa-bohu from Genesis 1:2 which is sometimes translated as ‘chaos’. I did not mean to suggest that, whatever that Hebrew term refers to, it is equivalent to what is referred to as chaos in mathematics. I simply intended to acknowledge that the word chaos carries connotations in theological contexts from that Biblical usage. Now, if you prefer a different translation for tohu wa-bohu, I’m happy to use something else if it will help clarify our conversation.
Thank you so much for your quick reply. I would like to approve of The New Living Translation like in “The Life Application Study Bible” (Tyndale):
“The earth was formless and empty (= a fluid lump of something muddy, not a lump of magma, with nothing on its surface. Not a lump of magma, because it was covered with “the waters”.)
In connection with the Wikipedia article “Logistic map”
To point out just a few of the self-contradictions of the notions of actual-infinity-th-step/stage fractals including the chaos-attractors-bases over a Moebius band or manifolds that are diffeomorphic to it:
1. Hausdorff dimensions fail to exist:
The plane-filling Peano-curve is said to have the 2.0000…-sharp Hausdorff dimension, which contradicts the very Hausdorff-dimensions-definition itself.
(1.5 The space-filling-curves notion holds another self-contradiction, as Lakoff and Nu~nes pointed out:
As those curves have finished filling out the spaces, to where do all the non-zero-width-ed gaps between the adjacent curve-parts [ = zero-width-ed lines] go/vanish?)
2. Self-similarity fails to exist:
As you know, the Sierpinski triangle and the Menger sponge have the parts/sub-spaces within them that are self-similar to their wholes. Namely, the magnification-functions f operating on those parts S generate S themselves; thus, f(S) = S; but that is the definition of an identity mapping; still, the magnifications are said to be no identity mapping.
Thank you for your reply. My answer is your last sentence itself. As also Dr. Michael Guillen pointed out in his book “Bridges to Infinity,” no rigorous mathematico-physical chaos-models can ever exist and have ever existed.
And as I have shown to you in one of my previous replies, there are counter-measure candidates for the catastrophe-theoretic models for the phenomena like the aphids-populations-fluctuations that usually get the tries (just vain/empty tries) at analyses by the logistic-mapping-models.
(I admit the book’s authors confess that their concrete catastrophe-models are still in the experimental stage, based on some insufficient data and have a big principle-wise problem that all the control-factors data are very hard to perfectly collect, but such models are the only theoretically-possible solutions.)
Let me ask you: have you not really seriously even looked over (= skimmed through) all my previous links? Namely, do you really intend to meet/face my criticisms squarely or not?
In case I forgot to link the ad for the above-mentioned book, here it is:
https://www.amazon.co.jp/Catastrophe-Theory-Aphid-Population-Dynamics/dp/3330018178
Thanks for answering my question; that is a helpful clarification. Yes, I am attempting to address your criticisms. Part of that is understanding them, which is why I am asking questions. Perhaps they are not the right questions, but they are the questions I think will help me understand, in terms that I am used to working with. Yes, I did look over the Twitter links you shared regarding your mathematical claims and proofs. Perhaps because of a lack of mathematical sophistication on my part, perhaps because I don’t read Japanese, or perhaps because I simply missed the point, I did not find the answer to my question there, which is why I asked it here.
Now that I believe I understand your objections to chaos theory and attractors, perhaps we can move on to the downstream claims. You seem to be drawing a connection from chaos theory to emergence to macroevolution. In what sense does macroevolution depend on chaos theory and emergence? As I recall, none of the mathematical models of macroevolution I linked above depended on chaos features like attractors or period doubling cascades.
Excuse me; regarding the relationship between macro-evolution and emergence and chaos dynamical systems, I have read only Japanese translations whose original English titles I have forgotten to take notes of. I would like you to go to the trouble of Net-searching for yourself/on your own. I am sure that you will find a plenty of info sources on it in the cyberspace. Again, I apologize for the inconvenience.
Excuse me, but this shows my frank review of your book:
https://www.amazon.co.jp/review/RS43GZQTTBCJP/ref=pe_1162082_181361352_cm_rv_eml_rv0_rv